黑视蛋白DNA适体可以通过改变分子钟的相位来调节哺乳动物昼夜节律的输入信号
调节生物节律钟针的黑视蛋白DNA 适体由丰桥技术科学大学和国家先进工业科学与技术研究所 (AIST) 团队开发。
DNA 适体可以特异性地与生物分子结合以改变其功能,因此可能使其成为理想的寡核苷酸疗法。我们筛选了 DNA 适体黑视蛋白 (OPN4),这是一种视网膜中的蓝光感光色素,在利用光信号重置中央时钟的昼夜节律相位方面发挥着关键作用。
首先,使用在细胞膜上表达黑视蛋白的细胞,通过八轮 Cell-SELEX 鉴定出 15 种黑视蛋白 DNA 适体 (Melapts)。随后,在稳定表达Period2 :ELuc 和黑视蛋白的成纤维细胞系中对每个 Melapt 进行功能分析,通过确定它们在蓝光刺激下重置哺乳动物昼夜节律相位的程度。在表达黑视蛋白的Period2:ELuc :胸苷激酶 (TK) : OPN4稳定成纤维细胞中监测了 24 小时内Period2 的节律表达。在主观黎明时分,观察到四个 Melapt 将其相位提前 >1.5 小时,而七个 Melapt 将其相位延迟 >2 小时。即使在没有光刺激的情况下,少数 Melapt 也导致相位偏移约 2 小时,这可能是因为 Melapt 只能部分影响相位偏移的输入信号。此外,利用Period1 ::luc 转基因 (Tg) 小鼠中少数 Melaps 诱发的相移,通过Period1节律表达来监测昼夜节律。
这些DNA适体可能具有在体内影响黑视蛋白的能力。总之,Melapts适体可以在体内和体外成功地调节哺乳动物昼夜节律的输入信号和移位相位(相位提前和相位延迟) 。
细节:
通过操纵黑视蛋白向中央时钟输入信号的能力来间接改善睡眠-觉醒周期,将具有社会和经济优势。
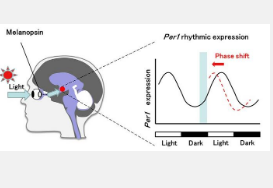
黑视蛋白是一种在视网膜神经节细胞中表达的光感受器蛋白,可吸收蓝光,最大吸光度为 477 nm。已知黑视蛋白在通过蓝光重置哺乳动物昼夜节律时钟相位和时钟基因(例如Period1,2 ( Per1,2 ))的节律表达中起着重要作用。分子昼夜节律时钟的相位由光刺激的时间和黑视蛋白光感受器对Per1的瞬时诱导重置并取决于这些时间(图 1)。最近,通过化学库化学筛选获得的黑视蛋白拮抗剂主要有助于延迟节律相位。
在本研究中,我们利用指数富集法 (Cell-SELEX) 的细胞系统性配体进化法来识别导致黑视蛋白改变昼夜节律相位的 DNA 适体 (单链 DNA;ssDNA)。总共分析了 15 种黑视蛋白适体 (Melapts 1–15),以评估它们在Per2 :ELuc:TK:Mel 稳定细胞中改变Per2 ::ELuc 生物发光振荡相位的能力,其中生物发光报告基因跟踪控制来自Pyrearinus termitilluminans的增强型绿色发光荧光素酶的Per2启动子区域,黑视蛋白在胸苷激酶 (TK) 启动子的控制下过表达。在这些稳定的成纤维细胞系中,信号通路被整合到成纤维细胞中,该细胞通过黑视蛋白模拟从视网膜到中央主时钟(视交叉上核或核:SCN)的信号通路(图2)。
DNA 适体是短的单链 RNA/DNA 分子,可以选择性地结合特定靶标、蛋白质、肽和其他分子,可用于临床切换靶分子的功能。这些适体的主要优点包括靶标特异性高、无免疫原性和易于合成。
在 15 个黑视蛋白 DNA 适体 (Melapts) 中,有 4 个 Melapts 诱导了相位提前,7 个 Melapts 诱导了 Per2 :: ELuc 细胞系中的昼夜节律延迟(分别延迟 >1.5 小时和 >2 小时)。即使在没有体外光刺激的情况下,少数 Melapts 也诱导了约 2 小时的相位偏移。
在光信号输入过程中,Melapt04 和 Melapt10 分别在 CT22 和 CT8 处诱导昼夜节律时钟相位提前或延迟约 3 小时。这表明 Melapt04 调节昼夜节律相位并促进入睡和觉醒,主要通过相位提前(图 3-5)。存在两种 Melaptes,它们以相同的方向提前和延迟相位移动,而不管光刺激的时间如何。然而,这三种 Melaptes 在黎明和黄昏时以相反的方向提前和延迟相位移动。因此,这些 Melapts 有望用于调节节律相位(图 6、7)。
免责声明:本答案或内容为用户上传,不代表本网观点。其原创性以及文中陈述文字和内容未经本站证实,对本文以及其中全部或者部分内容、文字的真实性、完整性、及时性本站不作任何保证或承诺,请读者仅作参考,并请自行核实相关内容。 如遇侵权请及时联系本站删除。